Pterosaur.net is a new website devoted to pterosaurs. Unfortunately, the authors of that site have provided much misinformation that can be clarified with available evidence. Below are several instances.
pterosaur.net An Introduction to Pterosaurs by Dave Hone
Hone claims that pterosaur wingspans ranged from around 50 cm to 13 meters.
Actually the lower range approached 13 cm for the tiniest adult pterosaur. Hatchlings would have been 1/8 as large (fly-sized) with a 1.6 cm wingspan.
Hone reported, "Pterosaurs are not dinosaurs (or birds for that matter), but close relatives of them. "
False. Phylogenetic analysis indicates that pterosaurs were more closely related to lizards and shared no traits with dinosaurs that were not found in greater abundance in fenestrasaur lepidosaurs, descendants of a sister to Huehuecuetzpalli.
Hone reported, "their exact origins are still uncertain."
False. Pterosaurs origins can be traced back to the origin of reptiles through diadectids, lepidosauriforms, basal lepidosaurs and fenestrasaurs such as Cosesaurus, Sharovipteryx and Longisquama.
Hone reported, "…and thus look quite different to their nearest relatives.
False. Ther are very few differences between pterosaurs and their nearest relatives. See Longisquama.
Hone reported, "…a small ‘tail vane’ flap on the end which may have also helped with steering.
False. Steering was accomplished by rolling the wings, as in airplanes, birds and bats.
Hone reported, "There are two major groups of pterosaurs …‘Rhamphorhynchoids’ were generally small, had relatively broad wings, short necks, short and robust skulls, a large rear wing membrane, were poor on the ground, and had (with one exception), long tails. Pterodactyloids were generally the reverse—big, with, narrow wings, long necks, long skulls (often with no teeth), a smaller rear wing, were good on the ground, and had short tails. >
False. There the two major clades in the pterosaur tree were dimorphodontoids and eudimorphodontoids. From among the eudimorphodontoids four clades independently developed smaller tails, longer necks and longer metacarpals. All pterosaurs and their ancestors were proficient on the ground (see below for more on this).
Hone reported, "...a special bone called the pteroid would have helped with steering."
False. The pteroid was a passive bone that formed part of the leading edge of the propatagium, a membrane spanning the gap between the wrist and shoulder.
Hone reported, "On the ground the ‘rhamphorhynchoids’ were probably pretty poor. Their large rear membrane would have shackled their hindlegs together making walking difficult, and the shape of their hips and upper legs meant that could only really sprawl and not walk upright.
False. There was no large rear membrane that shackled the hindlegs together. This model is based on a misinterpretation of the pterosaur Sordes. Actually the hind limbs were free of any sort of bind and operated like those of several living lizards capable of achieving high speeds while running bipedally.
Hone reported, "The pterodactyloids had no such problems, as can be seen by their extensive fossil record of footprints. They had split their rear membrane in two freeing the legs which were brought under the body to allow them to walk far more effectively than their predecessors and helped them diversify to exploit new environments and foods (see the terrestrial locomotion section).
False. No such split ever occurred. Uropatagia were all installed as original equipment. Pre-pterosaurs, including Cosesaurus, Sharovipteryx and Longisquama all had paired uropatagia that trailed each hind limb. The legs were not brought under the body in 'pterodactyloids.' They had wider sprawls relative to the earlier dimorphodontoids, which had a femoral "head" set at right angles to the shaft, similar to dinosaurs.
Hone reported, "Eggs were probably laid a few at a time and buried in nests."
False. Eggs were carried, one at a time within the mother, as demonstrated by Darwinopterus, a trait pterosaurs share with certain lizards. Archosaurs (birds and crocs) can't and don't retain eggs for long after fertilization.
pterosaur.net/anatomy Skeleton by Mike Habib
In Figure 2, Habib reported: "Potential membrane extents in pterosaurs. Current evidence suggests that a membrane completely free of the hindlimb was unlikely."
False. This statement was based on a misinterpretation of Sordes by Sharov (1971), Unwin and Bakhurina (1994) and Elgin, Hone and Frey (2011). Here and here are examples that show the hind limb was free of the wing membrane from mid thigh on down.
Habib reported: "Thanks to the discovery of wonderful fossils with soft tissue impressions from several sites around the world, we now know that at least some pterosaurs (probably most) had a wing that attached to the hind limbs (Fig. 2)."
As above.
Habib reported: "Currently, there are no specimens that clearly show an attachment free of the hindlimb.
False all the specimens show this. None do not. You can see more wing membranes at Ctenochasma gracile, Jeholopterus, Sordes, Rhamphorhychus, Pterodactylus, and here.
Habib reported: "Figure 4: Catapult mechanism in flexor tendons and fourth metacarpophalangeal joint, by Julia Molnar. The FDL tendon, pinned to the ground during stance phase, slides through the groove in metacarpal IV when the animal vaults, which releases stored energy suddenly. This greatly increases power output."
No manus imprints include digit IV, only digits I-III. Figure 4 reconstructs fingers I-III too small and dorsal to metacarpal IV when they should be larger and anterior to digit IV as shown here. Thus it would be impossible for metacarpal IV to be pinned to the ground and there would be no sudden release of stored energy.
Habib reported: "Based on reconstructions of tendon attachment points in pterosaurs, it is likely that some species (perhaps most) used a lock-and-release system on a flexor tendon cross the wing finger pivot - such a system is called a catapult mechanism (Figure 4)."
This was physically impossible. See above.
pterosaur.net/ecology Food, sex and over-excess: pterosaur palaeoecology in a nutshell by Mark Witton
Witton reported: "All three eggs share an extremely thin (30 μm) shell and were probably soft and leathery like the eggs of modern turtles and crocodiles."
Actually turtle and croc eggshells are much thicker. The pterosaur eggshell thickness is closest to that of lizards known to lay eggs in which the embryo was carried internally by the mother and hatchlings emerge minutes or days after expelled.
Witton reported: "This means that pterosaurs almost certainly had to bury their eggs underground or in piles of vegetation to prevent them drying out."
False. This is based on the false paradigm that pterosaurs were archosaurs related to birds and crocs. Actually mother pterosaurs carried their egg inside until just before hatching as demonstrated by Darwinopterus and several lizards.
Witton reported: "Furthermore, the reduced temperatures of these conditions probably extended the incubation time of pterosaur embryos to three months or so rather than the matter of weeks found in modern birds."
False. Inside a warm mother, development was accelerated.
Witton reported: "Size distributions clusters of single pterosaur species (notably those of the Jurassic form Rhamphorhynchus) suggest their growth rate was low: Rhamphorhynchus may have taken several years to achieve its full size (Fig. 1).
This false paradigm was based on Bennett 1995 who purported that distinct smaller species within Rhamphorhynchus were juveniles of larger species. The interval between hatchling and adult was described for Pterodaustro based on annular rings found in the bone of larger specimens (Chinsamy, Codorniu and Chiappe 2008). Likely the interval was longer in larger forms and shorter in smaller forms as in living birds and mammals.
Witton reported: "Fig. 1. The growth of Rhamphorhynchus muensteri. The tiny form on the left was once thought to be a distinct species from that on the right, but it’s now thought that they represent different growth stages of the same taxon. The smaller individual has a skull of 50 mm-or-so in length, while the bigger has a skull closer to 200 mm.
False. The smallers specimens were morphologically distinct from the larger ones. Start here to see the phylogenetic progression in size and morphology for Rhamphorhynchus.
Witton reported: "…frigate bird-like dip-feeding is unlikely for many pterosaurs because of their short, inflexible necks."
False. All dipping pterosaurs had a sufficiently long neck to dip feed. See here and here for samples.
Witton reported: "...no pterosaurs have the streamlined skulls or robust jaw joints of deep plunge divers."
False. Germanodactylus and all of its descendants have that sort of skull.
Witton reported: "Work on pterosaurs such as Tupuxuara, Germanodactylus, Pteranodon and Nyctosaurus has revealed that headcrests sprout very late in ontogeny, implying that their utility is strictly limited to adult life."
False. Small purported juvenile specimens also had large crests. Actually more primitive specimens had smaller crests.
Witton reported: "Research into the biometrics of several-hundred Pteranodon fossils shows that all known Pteranodon specimens are fully grown (or nearly so) and fall into two size classes, one sporting individuals approximately one third bigger than the other. Other than size, these groups differ in few anatomical details: the bigger Pteranodon have large headcrests and more pronounced extensions of the upper jaws over the lower, while smaller forms have comparatively broad pelvic canals."
False. This is based on the work of Bennett (1991, 2001) who did not employ a cladistic analysis which indicates smaller forms were more primitive (typically, but not universally). The large pelvis belonged to another taxon, Nyctosaurus, which is known for its large pelvic opening at any overall size. Distinct from Pteranodon pelves, the pelvis in Nyctosaurus separates the pubis and ischium. (illustration to come).
pterosaur.net/flight Life in the air by Mike Habib
Habib reported, "The more basal, paraphyletic “ramphorhynchoids” had a broad uropatagium that linked across the two hind limbs.
False. This is a misinterpretation of one specimen of Sordes by Sharov (1971), Unwin and Bakhurina (1994) and Elgin, Hone and Frey (2011). No specimens preserve this. All had separate uropatagia.
Habib reported, "...kicking a leg out would generate lift at an angle to the body, and thus make the animal roll or yaw to the side. In flying animals, a roll induces a turn, so pterodactyloids may have been quite maneuverable for their size.
False.
Actually a roll is induced at the wing tips, as in airplanes. The much smaller hind limbs would have acted like horizontal stabilizers or trim tabs on an airplane, stabilizing, not steering the pterosaur.
youtube/pterosaur launch animation
This video purports to show a pterosaur launching itself into the air using its forelimbs to achieve enough "air time" to unfold and flap its wings, like a vampire bat. Unfortunately we know of no launch sequence footprints to confirm this. Some pterosaurs, such as MPUM 6009, had such short forelimbs and such long hindlimbs that a crouching launch was unlikely.
The narrow margin of error needed for the pterosaur to unfold its (sometimes huge) wings thousands of times during a lifetime without ever striking the ground seems to be improbable. Moreover, the power and coordination needed to complete the vault and extend the wings in this fly-or-crash scenario gives little room for a learning curve in young pterosaurs or in the evolution of such a maneuver from predecessors starting from a standard hind limb leap.
While we're on the subject of pterosaur launching, here are three GIF movies showing 1) a successful forelimb launch, 2) an unsuccessful forelimb launch and 3) a successful bird-like hindlimb and wingflap launch with headwind. Without a headwind a running start, as in the albatross (see below), would have been necessary. Animation is always biased, but here the other sides of the question of pterosaur launch are at least offered. In the hind limb launch (3) three complete wing beats occurred in the same time as less than one beat in the upper two. It also demonstrates that pterosaurs only had to raise their wings to become bipedal while maintaing their center of balance over their toes.

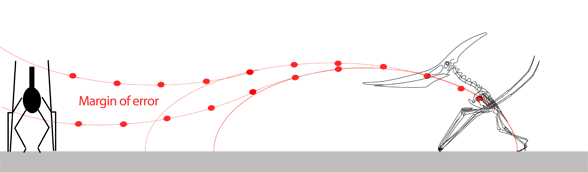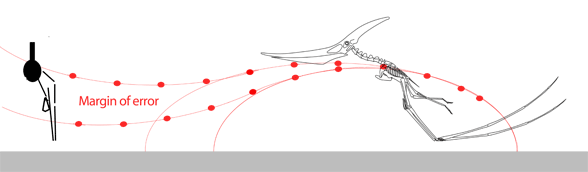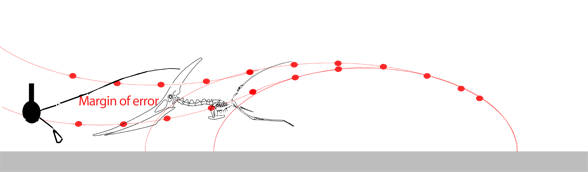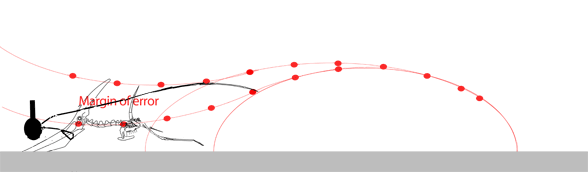

Habib (2009) wrote: "…a quadrupedal
launch would allow much greater leaping forces by including
the musculature of the flight apparatus, such as the coracobrachialis and pectoralis muscles, and thus produce high launch speeds."
Unfortunately, Habib (2009) did not report a specific launch speed or range. As lizards capable of high speed bipedal locomotion (see below), a grounded pterosaur in a no-wind condition could have run to attain sufficient airspeed, just like an albatross (see below). However, a headwind provided by a sea breeze would have provided sufficient airspeed to produced lift without the need to run.
With regard to muscle size, the thighs extend the length of the ilium, which is comparable to the length of the pectoral girdle or humerus.
Pterosaurs descended from hind limb leapers, such as Sharovipteryx, Longisquama and the basal pterosaur, MPUM 6009, all of which were speedy runners, able to balance bipedally over their toes. The thighs and femora were well designed for pushing off. Wings were built to flap, cling and swing back and forth during quadrupedal excursions. No propulsion vectors were provided by these forelimbs, only support (see GIF movie below). There is no olecranon process, as there is in the Desmodus, the vampire bat, to provided mechanical leverage for rapid extension under weight.
When kangaroos make their initial hop (before reaching top speed), their toes reach the heights of their ankles. It seems unlikely then that a pterosaur with much less leaping ability in the forelimbs, could do any better without wings to help.
| From a standing start in a no wind situation, several albatross perform a running take-off in the video at right. Note the amplitude and frequency of the wing beats. While a maginitude smaller than Pteranodon, the proportions of the bird and pterosaur were similar. The frequency of wingbeats would have been reduced due to the mass and momentum differences in the wings. Larger means slower. With a headwind, an albatross can take off from a standing start. Without a headwind, Pteranodon might have needed a running start. |
 |
| As lizards with sprawling femora (see below), pterosaurs would have run iike lizards, perhaps during take-off. At right, in this video from the Bruce Jayne lab in Cincinnati, Ohio, a zebra-tailed lizard, Callisaurus draconoides, is shown running quadrupedally and later bipedally (Irschick and Jayne 1999). Note the heels never touch the ground. Note the incredble speeds: 5 m/sec or 11 mph. Note that footfalls occur every three body lengths. What could Pteranodon do? Due to its size, the frequency might have been reduced, but the speeds may have increased. We'll have to wait for running/takeoff footprints. |
 |
pterosaur.net/terrestrial locomotion Terrestrial locomotion in pterosaurs by Mark Witton
Witton reported, "First and foremost, Pteraichnus and its ichno-kin demonstrate to us that pterosaurs walked quadrupedally on digitigrade forelimbs and plantigrade hindlimbs.
Only partly true. Certain pterosaurs walked this way while beachcombing and footprints demonstrate this. Other pterosaurs did not walk quadrupedally as described by Peters (2000).
Witton reported, "Only the first three fingers regularly touch the floor as the flight finger was apparently folded and stowed alongside the body when grounded, occasionally leaving a fourth digit trace in the manus print.
False. A fourth digit trace has never been found.
Witton reported, "...pterosaur manus prints fall almost directly anterior to the pes prints with only slightly more pronounced displacement from the midline. Hence, reconstructing a pterosaur standing on such trackways shows that both the hindlimb and forelimb are not splayed anywhere near as much as suggested by earlier workers.
False. Due to the right angle of the knee, a splayed femur returns the ankle close to the midline (see illustration below). Furthermore, manus prints are always somewhat lateral to the pes print and often posterior to it, as shown below.
| Animation of Pterodactylus no. 21 walking in Crayssac tracks. Note that the weight was born principally by the hind limbs with the front limbs acting as ski-poles or stabilizers. During push-off the manus was behind the shoulder giving thrust to the entire arm. That doesn't happen in competing configurations. Note also the more erect pose here. Competing configurations adopt a more horizontal pose. |
|
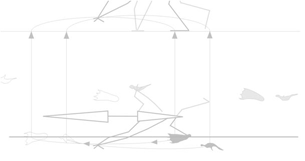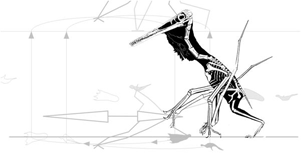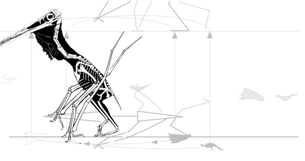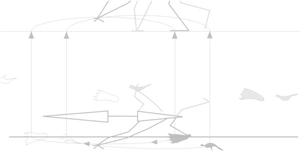 |
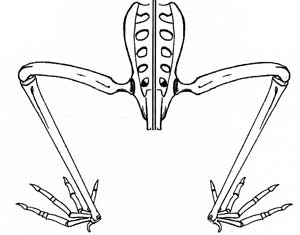
Witton reported, "...the limbs of pterosaurs are held parasagitally (Fig. 4), a feat achieved by retracting the humerus posteriorly and ventrally in the glenoid and the highly mobile joints of the acetabulum and knee joints. This gives pterosaurs a relatively efficient stance with their bodyweight carried on near-columnar limbs rather than the energy-sapping stance of sprawling reptiles and amphibians.
False. The knees were bent. The reconstruction in Witton's Fig. 4 keeps the manus in front of the shoulder and elbow throughout the step cycle, so there is no thrust generated from the forelimb in that configuration.
| Pelvis and hind limbs of Pteranodon. Here the feet were close to the midline and closer if the knees were lower in relation to the hip joint. These are lizard-like hips, more splayed than those of Dimorphodon, for instance. |
 |
Witton reported, "...exceptionally well preserved basal pterosaurs appear to have a single, broad cruropatagia that extends fully between the hindlimbs and pelvis with its ventral margin supported by the elongated fifth toes of the pedes.
False. Witton is referring to misidentified torn wing material in Sordes, which can be seen here.
Witton reported, "Basal pterosaurs typically have deepened, highly recurved manual and pedal claws with comparatively large flexor tubercles compared to the relatively slender claws of pterodactyloids.
False: See SMNK PAL 3830.
|
