|
|
 |
|
Pterosaur wing origins
It has been said that pterosaurs appeared suddenly in the fossil record with no apparent precursors. Unfortunately, this is the belief of those who still believe pterosaurs are archosaurs. No series of archosaurs has ever been put forth that demonstrates a gradual evolution of pterosaurian characters, hence the paradigm mentioned above.
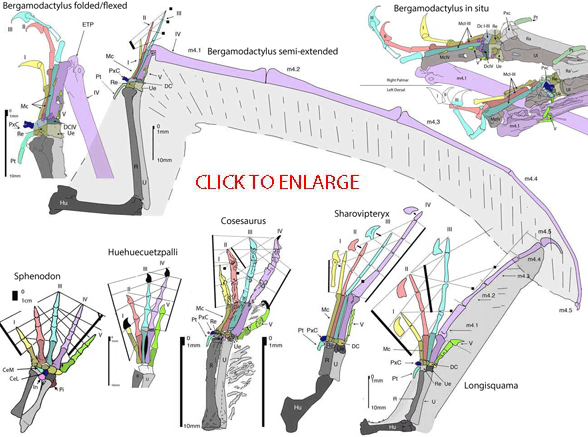
Lepdiosaurian fenestrasaurs (Peters 2000, 2009) do demonstrate a gradual evolution of pterosaurian characters, no matter which characters one chooses. Here the forelimb is transformed into a wing using a series of lepidosaurs.
1.7 Illustrates the basal lepidosaur, Sphenodon. Digit 3 extends further than digit 4. Metacarpal 3 is longer than 4. All the carpals are present including two centralia. The medial centralia is elongated.
1.1 Illustrates the basal squamate Huehuecuetzpalli. Here the carpals are poorly ossified, even in the adult. As a result the distal radius and ulna touch because the intermedium is absent. Metacarpal 4 is as long as 3 and digit 4 extends further than digit 3. Digit 5 is somewhat shorter.
1.2 Illustrates the basal fenestrasaur, Cosesaurus. Here the radius and ulna have little to no space between them (compare to 1.7) and the carpals are well ossified. Thus supination and pronation were dramatically reduced. Note the two centralia are now on the medial side of the carpus where they take on new identities as the pteroid and preaxial carpal (Peters 2009). Only fenestrasaurs, including pterosaurs, have this arrangement. The digits are robust and digit 5 is further reduced. Extradermal tendrils trail the ulna and finger IV, probably for decoration. Cosesaurus was occasionally bipedal and likely flapped its arms due to the presence of a pterosaur-like pectoral girdle.
1.3 Illustrates Sharovipteryx. This fenestrasaur reduced its forelimbs and elongated its hind limbs so much that the forelimbs never touched the ground during locomotion. Here the stunted radius and ulna are shorter than the hand. The gracile fingers are much longer laterally with deep claws. Digit 5 is further reduced. The extended neck skin acted like strakes. The flapping forelimbs acted like canards and probably provided some propulsion for this hindlimb glider.
1.4 Illustrates Longisquama. This fenestrasaur has a pterosaur-like wing membrane trailing the forelimb, which was relatively much larger, but otherwise similar to Sharovipteryx. The axial rotation of metacarpal IV has begun because digit 4 no longer had continuous flex lines with digits 3, 2 and 1. This rotation enabled digit 4 to flex in the plane of the wing for wingfinger folding. Such a wing would not have provided enough lift for flight, but it could have extended glides by providing thrust.
1.5 and 1.6 Ilustrate the derived fenestrasaur and basal pterosaur, MPUM 6009. Here the hand is much smaller, but the wing finger is hyper-elongated. The fiber-embedded wing membrane (poorly preserved in the original fossil) is large enough to provide thrust and lift for flight. While Longisquama used its wings primarily to ward off rivals and entice mates, MPUM 6009 could have used its wings to fly from place to place in powered flight. Later pterosaurs modified the wing elements in various relatively minor ways. Only one pterosaur, SOS 2428, reduced its wings and became flightless. Note digit 5 is not gone. It is a vestige. More examples of manual digit 5 in pterosaurs are here. The wing ungual is also not gone. However it does become disarticulated on most pterosaur specimens. Note that a trochlear joint for the reception of a wing ungual always tips the fourth wing phalanx. Examples that appear pointed are the result of a partially buried tip that needs further preparation.
A recent paper on pterosaur wing shape (Elgin, Hone and Frey 2010) purported to show that all pterosaur wing membranes attached at the tibia or ankle (see below). Unfortunately, all of the examples presented demonstrated the opposite, that the wing was stretched between the wing finger and elbow with a small fuselage fillet inboard as shown above and here. Elgin, Hone and Frey (2010) discarded one of the best examples of the narrow chord model, the Zittel wing. They considered other narrow chord wings as examples of "membrane shrinkage," presumeably due to desiccation. That is an invention enforced my misunderstanding examples of wing membrane tearing and folding. Wing membranes don't shrink. They are embedded with fibers, not water. |
|
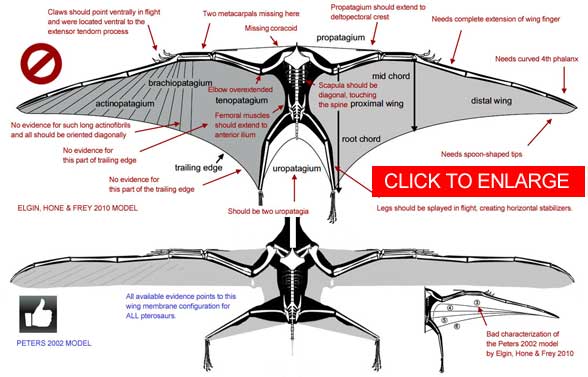 |
| Click on the above image based on the Elgin, Hone & Frey (2011) report on pterosaur wing shape to enlarge it. |
|
 |
Just to clear the air, David Hone in his blog Archosaurmusings.com published the above image of the distal phalanx of an unidentified pterosaur wing on June 29, 2010.
Hone reported phalanx IV tapers to a point, which it does not. Actually it tapers to a trochlear interphalangeal joint.
Hone wrote, "Finally it often has a slight pathology (as seen here) of the tip curving to point slightly (or occasionally, very) posteriorly. [And more here]" Actually this is the fifth phalanx, the ungual or claw, which is traditionally considered missing in pterosaurs, but, as you can see, it is very much present in all pterosaurs. Often it is disarticulated, hence its apparent rarity.
The above image is used under the protection of Section 107 of US copyright law which lists various purposes for which the reproduction of a particular work may be considered fair usage, such as criticism, comment, news reporting, teaching, scholarship, and research. In this case most of the categories in this list are in play. |
|
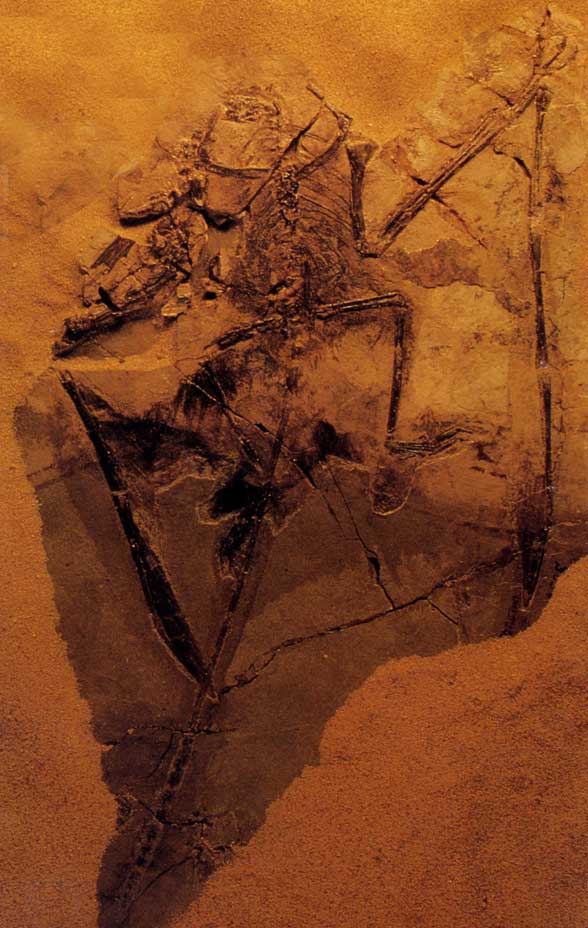 |
The holotype of Sordes pilosus PIN 2585/3. Roll over the image with your mouse pointer to see how the membrane
and bones have shifted after death.
The Sordes problem. Much ado has been made about the Sordes holotype with regard to pterosaur wing shape. On the left it appears as though some membrane makes a straight line toward the left ankle. Nevermind that the line keeps going off to the right. Earlier (Peters 2002) I thought that a geological fault was "at faul"t for creating such a straight line on both sides of the leg. That's not the entire answer. The key to solving this problem was making sure all the other elements were present. Most were.
However the antebrachium (radius + ulna) was missing on the left...or so it would seem. The first wing phalanx was bent in unnaturally and everyone thought the skull was damaged. Apparently it is largely missing too. In its plac, or on top of it, is the left hand with metacarpal IV separated from I-III and the wrist unnaturally close to the elbow. The radius and ulna have drifted posteriorly where one forms the straight line from the wing to the ankle and the other croses it. That's why Sordes seems to have a different wing shape than all the other pterosaurs, which you can see a sampling of here and here.
Note that the right wing is intact and extends to the elbow, not the ankle. The right iinner wing was torn from its mastings, folding much like one might expect a uropatagium to fold. Everyone thought Sordes had uropatagia much deeper than any other pterosaur. In fact the uropatagia were quite similar to those of other pterosaurs, as shown above in red. Other tissues were caught on the feet and ankles in the same current or event that dislodged the left antebrachium before burial.
This is also a fine example of wing ungual retention (see above).
You can see more wing membranes at Ctenochasma gracile, Jeholopterus, Sordes, Rhamphorhychus, Pterodactylus, and here. |
|
![The Zittel Wing [Rhamphorhynchus] insitu](images/lepidosauromorpha/diadectidae/lepidosauriformes/fenestrasauria/pterosauria/zittelwing1.jpg) |
The Zittel Wing, belonging to a Rhamphorhynchus specimen, displays the true shape of the wing membrane, stretched essentially between the elbow and wingtip with a small fuselage fillet extending posteriorly toward mid-thigh. The inner membrane was not reduced by shrinkage (contra Elgin, Hone and Frey 2010) as no other part of the wing membrane was reduced. Instead embedded fibers prevented shrinkage. Roll over the image to see the extent of the wing membrane. |
|
 |
No pterosaur ichnites currenly known preserve a pterosaur launch, but Habib (2010) proposed a forelimb launch sequence. Here a more traditional hind limb launch is favored. As in the albatross, a standing take-off may only have been possible into a headwind. |
|
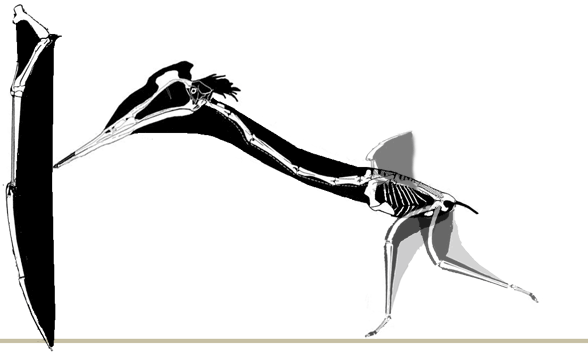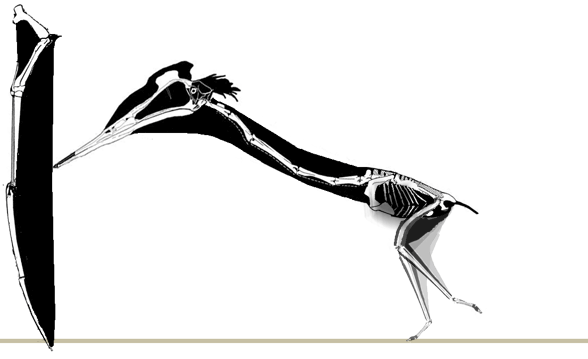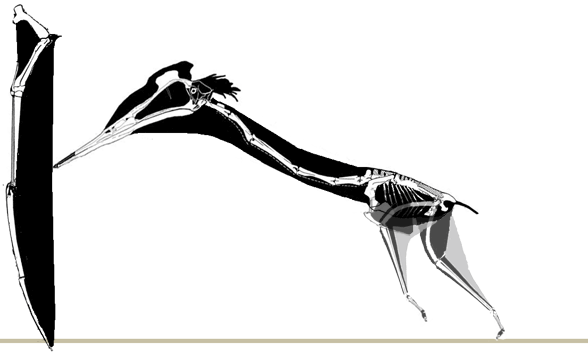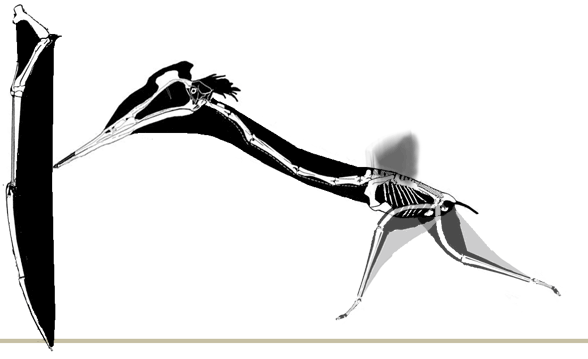 |
A running take-off, again in the pattern of the albatross, creates its own headwind. This type of running also follows the pattern of bipedal lizards shown here in which the heels never touch the ground and the trackway is narrower. |
|
 |
GIF animation of a pterosaur, Pterodactylus longicollum, matched to Crayssac tracks demonstrating a landing sequence (Mazin, Billon-Bruyat and Padian 2009). |
|
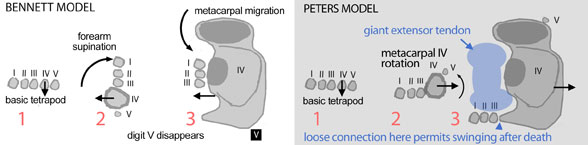 |
Two reconstructions and evolutionary models have been proposed for the evolution and configuration of pterosaur wings. In the Bennett model (left) the primitive five-fingered hand supinates presenting the palms forward. Then digit 5 disappears and digit 4 enlarges while digits 1-3 migrate to the anterior (former ventral) side of metacarpal 4. In the Bennett model the wing finger hyper extends to fold upon the metacarpus. The former flexors extend the wing and the extensors flex the wing finger. No fossils preserve this model. There is no space for the extensor tendon.
In the Peters model only metacarpal 4 rotates posteriorly. Digit 5 becomes a vestige. Flexors remain flexors. The giant extensor tendon was dorsal to metacarpals 1-3. |
|
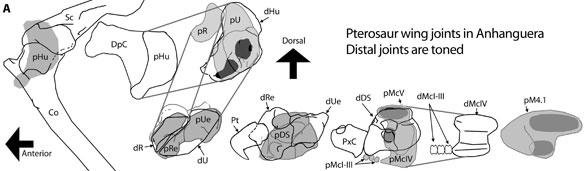 |
| Here the various joints in the proximal pterosaur wing are shown in distal view. |
|
|
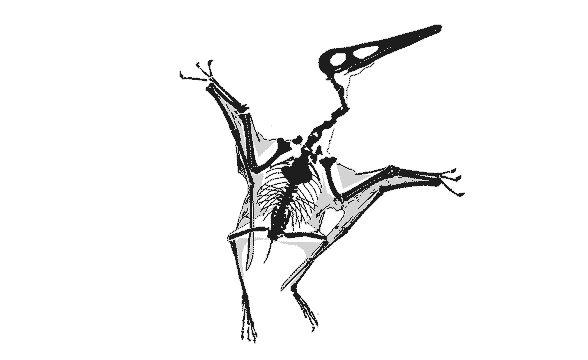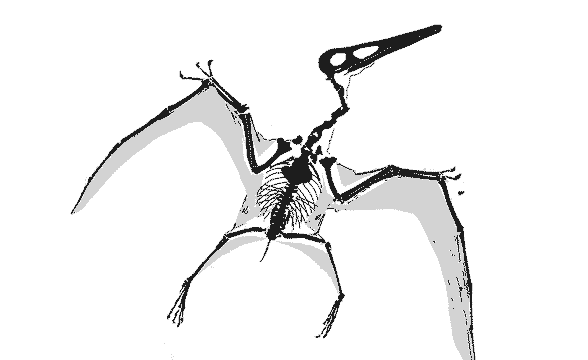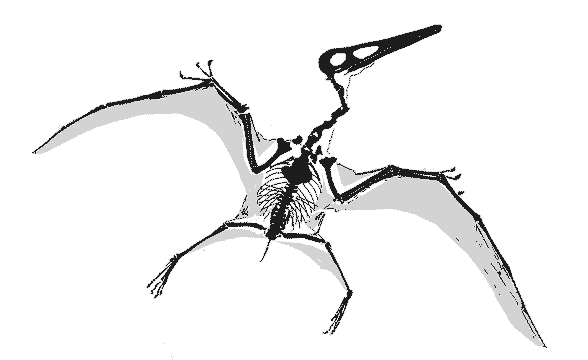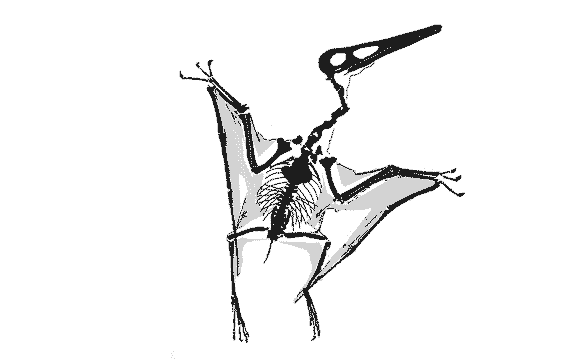 |
GIF animation of the Vienna Pterodactylusdemonstrating wing extension as reported by Peters (2002). There is no membrane extension possible for the wing membrane posterior to the elbow (the fuselage fillet) to create the deep chord, bat-like wing membrane proposed by Elgin, Hone and Frey (2010) because there is no membrane material there. Neither do the wing membranes attach at the ankle, contra the traditional and imaginary model supported by Elgin, Hone and Frey (2010). There is no "shrinking" of the wing membrane in this fossil either. Wing membranes don't shrink. They tear and fold only due to the imbricating aktinofibrils. This model permits the wing membrane to fold, like a Japanese fan, into the smallest possible area when retracted/folded. A deep chord membrane would droop like a blanket, as demonstrated in various movies improperly featuring deep-chord wing membrane pterosaurs, including those in Jurassic Park III.
Only at full extension do the wings have an aerodynamic shape. If pterosaurs folded their wings in flight, like some birds do, lift would be lost. Also note the relative placements of the elbow, pteroid, wrist and hind limbs. Yes, the hind limbs would have acted like the horizontal stabilizers on a standard airplane, a trait inherited from ancestors as far back as Sharovipteryx. The hind limbs were capable of lateral extension, like those of lizards, not dinosaurs and Scleromochlus. |
|
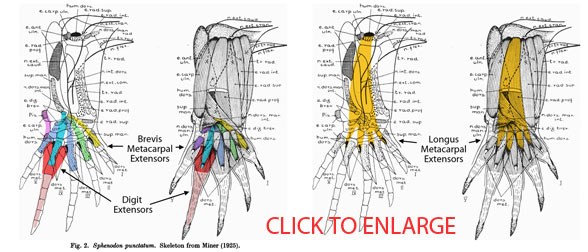 |
Extensor muscles and tendons in Sphenodon. Click to enlarge. Compare to hypothetical extensor muscles in Cosesaurus and a basal pterosaur (below).
|
|
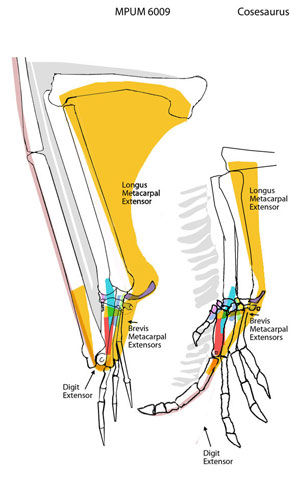
Roll over image to view extension and flesion in the pterosaur wing and cosesaur finger. He are hypothetical extensor muscles in Cosesaurus and a basal pterosaur, the Milan specimen, MPUM 6009. Compared to Sphenodon (above) the metacarpals are appressed and metacarpal 4 is axially rotated to facilitate wing folding in the plane of the metacarpus. Automatic opening of the wing finger with elbow extension (Prondvai and Hone 2009) had to be moderated because pterosaurs also used their forelimbs for walking, which involved elbow extension.
Here the pteroid extends anteriorly when the propatagium (extensor digitorum longus) is relaxed, but is pulled medially when the propatagium is pulled taut by the humerus. When the pteroid is pulled medially, a tendon from the pteroid extends the wing finger |
|
 |
|
| |
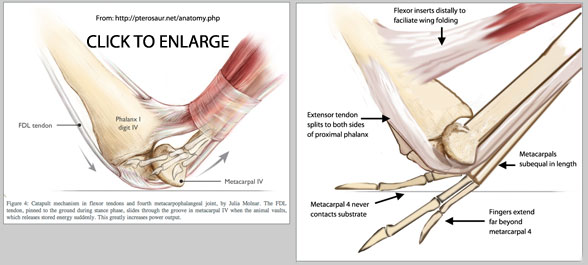 |
Another look at the so-called catapult mechanism during pterosaur take-off. Dr. Mike Habib (2008) proposed a fore wing launch method for pterosaur take-off which was illustrated by Julia Molnar online here (and see above, left). A catapult mechanism (as in grasshopper hind legs) was specified to increase the power of the launch. Unfortunately the illustration by Molnar includes several errors which preclude the possibility of a catapult mechanism in pterosaurs. Click to enlarge.
Molnar illustrated metacarpals 1-3 way too short. On the right the correct length is indicated. All four metacarpals were actually the same length. That means fingers 1-3 extended beyond the metacarpus. Pterosaur manus ichnites only impress digits 1-3. No trace of digit 4 ever impressed. Thus there was no pinching of the extensor tendon beneath the weight of the pterosaur to load the catapult prior to launching.
Furthermore, as in lizards, the long metacarpal extensor actually split prior to the knuckles to insert on the medial and lateral sides of the proximal phalanges. They did not extend over the knuckle. Much smaller muscles and tendons, the digit extensors (see above and here) extended over the knuckles.
Finally the flexors had to insert further distally than Molnar illustrates. In the Molnar illustration the flexors reach the limit of their ability to flex at 90 degrees. By inserting the tendon further distally flexion can continue until the wing finger is completely folded (see sequence here). |
|
|
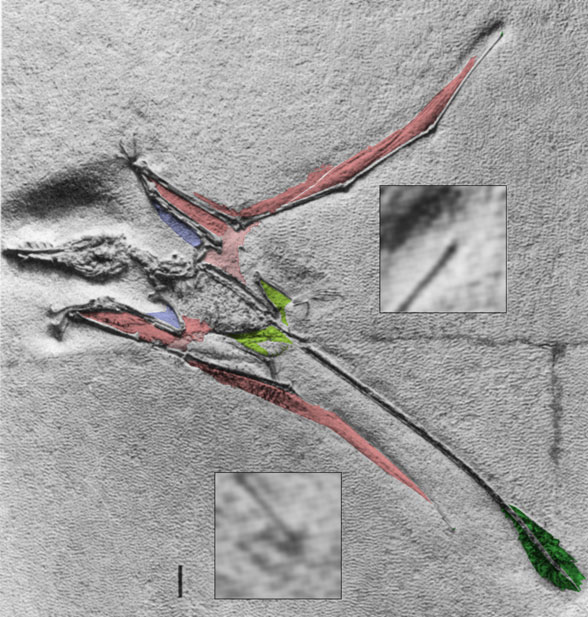 |
Rhamphorhynchus sp. BSP 1938 I 503a, No. 11 in Wellnhofer 1975. Click to enlarge and see other samples. Elgin, Hone and Frey (2011) claimed in this example that the wings had experienced "membrane shrinkage." This is not true. They are merely twisted in this case, twisted to such an extent that the membrane is anterior to the wing finger. |
|
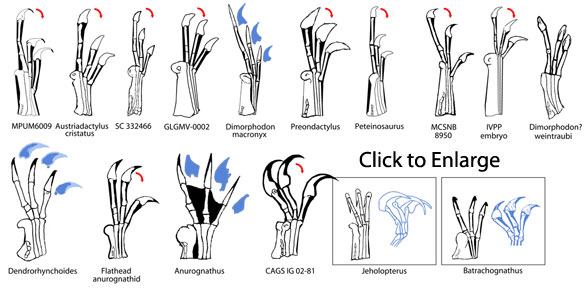 |
Pterosaur fingers. Click to enlarge and read the caption. |
|
| |
Bennett SC 2008. Morphological evolution of the forelimb of pterosaurs: myology and function. Pp. 127–141 in E Buffetaut and DWE Hone eds., Flugsaurier: pterosaur papers in honour of Peter Wellnhofer. Zitteliana, B28.
Elgin RA, Hone DWE and Frey E 2011. The extent of the pterosaur flight membrane. Acta Palaeontologica Polonica 56 (1), 2011: 99-111. doi: 10.4202/app.2009.0145
Habib M 2008. Comparative evidence for quadrupedal launch in pterosaurs. Pp. 161-168 in Buffetaut E, and DWE Hone, eds. Wellnhofer Pterosaur Meeting: Zitteliana B28
Mazin J-M, Billon-Bruyat J-P and Padian K 2009. First record of a pterosaur landing trackway. Proceedings of the Royal Society B doi: 10.1098/rspb.2009.1161 online paper
Peters D 1995. Wing shape in pterosaurs. Nature 374, 315-316.
Peters D 2002. A New Model for the Evolution of the Pterosaur Wing – with a twist. - Historical Biology 15: 277–301.
Peters D 2009. A reinterpretation of pteroid articulation in pterosaurs. Journal of Vertebrate Paleontology 29:1327-1330.
Prondvai E and Hone DWE 2009. New models for the wing extension in pterosaurs. Historical Biology DOI: 10.1080/08912960902859334
Sharov AG 1971. New flying reptiles fro the Mesozoic of Kazakhstan and Kirghizia. Trudy of the Paleontological Institute, Akademia Nauk, USSR, Moscow, 130: 104–113 [in Russian].
Unwin DM and Bakhurina NN 1994. Sordes pilosus and the nature of the pterosaur flight apparatus. Nature 371: 62-64.
Zittel KA 1882. Über Flugsaurier aus dem lithographischen Schiefer Bayerns. Palaeontographica 29: 7-80. |
|
|
|
|
|
